 Open Access
Open Access Abstract
Stem cell has great therapeutic potentials as it proliferates indefinitely, as well as gives rise to other cell type in our body. However, human pluripotent stem cell (hPSC) technology faces some obstacles associated with tumorigenicity and telomere shortening. And the risk of tumorigenicity of hPSC upon transplantation is one of the major hurdles, which must be overcome before hPSC based clinical practices. Interestingly, human amniotic stem cell (hASC) showed promising results by bypassing from the drawback. But, the important question is how hASC fully or partially escape from the progression of teratoma in severe combined immunodeficiency (SCID) mice, which remains unravel. It is decisive to comprehend the molecular mechanisms responsible for that of teratogenic and this non-teratogenic effect. Evidently, teratoma represents a critical line between stem cells, differentiation and tumorigenicis. And hASC typifies as a transitional stage between human embryonic stem cell (hESC) and adult stem cell. Hence, the study of hASC as a comparative model to reverse teratoma/teratocarcinoma formation and stem cell pluripotency to deciphering the molecular signals pathways promoting teratoma by hESC/hiPSC but not by hASC. Overall, learning the mechanisms of teratoma formation reversely by inducing in hASC could provide the knowledge that improves the tissue reconstitution potential from hESC and human induced PSC.
Human Amniotic Stem Cell
Stem cells research and progress open a plenty of hope, but halting clinical applications due to the chance of rejection by the recipient’s immune system and the transgene based induction of pluripotency. However, the multipotenthASC, future of regenerative medicine, is non-tumorigenic and immune privileged cell, maintains telomere length in comparing to hESC and hiPSC in culture dish. Recently it has been identified that C-Kit+hASC shares 82% transcriptome identity with hESC. It also expresses stem cell factors of Oct3/4, Sox2, Klf4, c-Myc, Nanog and Lin28 (Shown in Figure 1 ), it can replicate as undifferentiated cells Koike et al., 2014 Moschidou et al.,2013 . Human ASC represents an intermediate stage between hESC and iPSC/adult stem cell and are thought to be closer to PSC than bone marrow stem cell Si et al.,2015 . Unlike hPSC, however, hASC do not form a teratoma in SCID mice De Coppi et al., 2007 Ilancheran et al., 2007 . Indeed, an understanding the mechanisms whereby hASC does notform tumors, despite possessing similar molecular machinery to hESC and hiPSC, may be useful in directing lineage specific and tumor suppressing differentiation of hPSC. Generally, hASC isolated from amniotic fluid contains a heterogeneous population of differentiated and undifferentiated, and progenitors of some cells those are contributed mainly from embryonic/extraembryonic origin Scherjon et al., 2003 . Notably, hASC expresses immuno-suppressive factors CD59 and HLA-G but not HLA class II Li et al., 2014 . The hASC exhibits multilineage potential and can differentiate into the three germ layers of ectoderm, mesoderm and endoderm. Recently, CD117+/CD44+ hAFSC has shown to differentiate into oocyte-like cells in vitro Yu et al., 2014 .
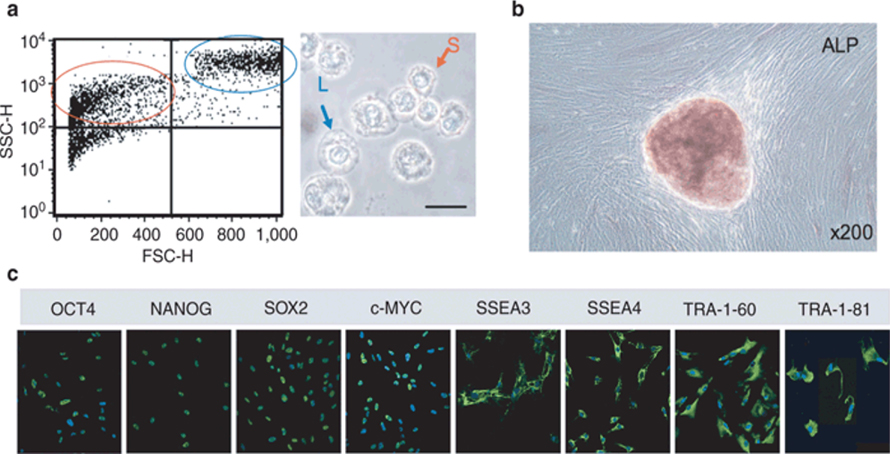
Figure 1. Heterogeneity of the first-trimester hASC population
Teratoma/Teratocarcinoma
Teratoma/Teratocarcinomarepresent a critical interface between stem cells, differentiation and tumorigenesis. It expresses the network of core transcriptional factors to promote the undifferentiated state. Simply, it is the counterpart of ESC but tumorigenic Fuchs et al., 2014 . On one hand, teratoma has the potency to self-renew in the absence of LIF or of a feeder layer, and differentiated into all three germ layers. Cultures of pluripotent teratoma cells are heterogeneous in nature. In 1974, Martin and Evans reported that the presence of two types of cell named E and C type in SIKR (a type of teratoma) culture. Together in culture, C and E sub clones were found to be tumorigenic and pluripotent. When they isolated both sub clones separately and tested for tumorigenicity, only C type showed positive result. Morphologically, the non-tumorigenic E type clone was fibroblastic, which was distinctly not embryonal carcinoma (EC) Martin and Evans, 1974 . On the other hand, hASC is a heterogeneous population of cells, non-tumorigenic, morphologically fibroblastic, and can self-renew in the absence of LIF or of a feeder layer. There has not, however, as yet any detailed comparative description of the characteristics in vitro of these cells. It would be motivating to confirm whether hASC could induce transformation effects similar as the E type transform to teratogenic in association with C type clone. Double immunohistochemical staining confirmed that hASC simultaneously expressed epithelial cell markers with mesenchymal ones Davydova, 2010 , and this sort of co-expression has been noted in circulating tumor cells or metastasis associated EMT/MET Armstrong et al., 2011 Papadaki et al., 2014 . Another fascinating study reported that valproic acid (VPA) supplemented hESC medium induced reprogrammed c-Kit+ hASCs are capable of forming embryoid bodies (EBs) in vitro and teratomas in vivo (Shown in Figure 2 ) Moschidou et al., 2012 . Hence it may be speculated that common factors may present in ESC and iPSC but absent or down regulated in hASC which inhibit teratoma/teratocarcinoma formation, and that or those acts as inhibitor/inducer/regulator individually or together with others, which could induce transformation.
Figure 2. Transplantation of hASCs into SCID mouse testes (Left figure)
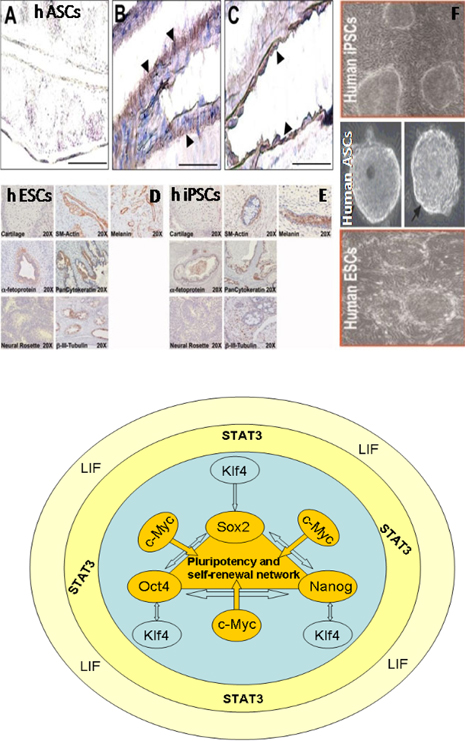
A) Absence of immunostaining for KRT7 in a tissue section taken from a SCID mouse testis injected with saline. B) A cell monolayer composed of KRT7-positive cells surrounding a murine blood/lymph vessel indicated by arrowheads. C) Serial section with the blood/lymph vessel shown in B identified by positive immunostaining for murine endothelial cell antigen PECAM1 (arrowheads). D & E) Transplantation of hESCs and hiPSCs into SCID mouse testes, and F) Embryoid body formation. Bars = 100 μm. Right figure shows the core pluripotency and self-renewal factors interaction. Figure adapted from Moschidou et al, 2012.
However, unlike most cancerous tumors, the polyclonal teratoma originates from a group of apparently normal cells instead of from a single abnormal cell Blum and Benvenisty, 2008 . The key regulatory stem cell self-renewal marker Oct4, Nanog, and Sox2 found to be down regulated by genistein in embryonic carcinoma Regenbrecht et al., 2008 . But, GAL and Oct4 were up- regulated in TGCT validated diagnostic markers for undifferentiated tumor cells that means in teratoma. Previously it was reported that genistein treatment induced Gadd5A/G in prostate cancer cell line Oki et al., 2005 . So, it could be speculated that genistein indirectly down regulates Oct4 possibly via the up regulation of Gadd5A/G. Again, temporary over-expression of miR27 directs to decrease the levels of Oct4 in hEC including DNA methylation Fuchs et al., 2014 . In gastric cancer cells mir27 activates metastasis and induce the epithelial to mesenchymal transition (EMT) Sun et al., 2003 . Normally hEC cells are detected by expression of Oct4 and Nanog that thought to be activated by NR5A2 Sung et al.,2012 . Nevertheless, as Oct4 is not a direct target of miR-27, and that of Oct4 and miR-27 form an indirect negative feedback loop where another unknown player may be involved. In miR-27 directed hESC developmental fate determinant genes of Tgfß- and the ßsignalling are noticed to be significantly up-regulated. Mechanistically, miR-27 negatively regulates the expression of the pluripotency-associated Activin/Nodal axis (Smad2/3) of the Tgf-ßsignalling pathway by targeting receptor Acvr2A, TgfßR1 (Alk5) and downstream effector Smad2 Fuchs et al., 2014 . And in the downstream the integrator Runx1 is a target for miR-27 to promote granulocyte differentiation. Micro RNA27 based inhibition of APC of Wnt signaling was reported in mesenchymal cells. But the complete study of the effect of oncomirs and anti-oncomirs yet to do. Retinoic acid (RA) treated Ntera2 EC showed to express Wnt-13 and differentiated into neurons during the early phases and later followed non neural pathways upon addition of hexamethylenebisacetamide (HMBA) in culture media Wakeman et al., 1998 . Mutated Lrrc50 and BMPR1bb (Alk6) are specifically linked to seminoma predisposition in humans. Bmpr1bb is significantly down regulated in response to RA treatment during patterning Chen and Amatruda, 2013 . Lrrc50 is solely as a ciliary motor protein has emerged as a signaling center for Tgfß and Wnt pathway and genes related to DNA methylation and imprinting Basten et al., 2013 . So, it seems like there is a role for Tgfß and Wnt signaling in controlling the fate decision in teratoma formation. We propose here an incomplete teratoma and pluripotency interlinked network that shown in Figure 3 .
Figure 3.
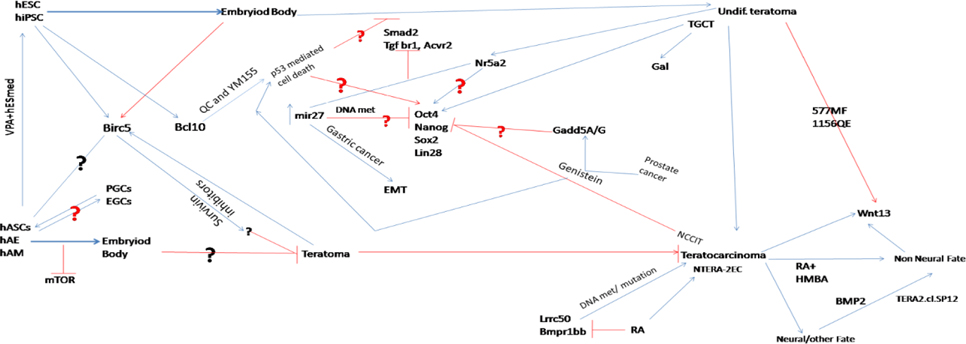
Tentative teratoma and pluripotency interlinked network for iPSC/hASC/ESC. The interlinked roles of the players remain to reveal.
Escaping tumor formation by stem cell
It was recently found that the ablation of survivin, both genetically and pharmacologically, stimulated apoptosis in cultured teratomas. Two genes namely Birc5 and Bcl10 identified to associate with embryonic carcinoma Blum et al., 2009 Lee et al., 2013 . Transcriptome analysis revealed that Birc5 (survivin) is highly expressed in hESC and teratomas but not in embryoid bodies. Taxol with purvalanol A (survivin inhibitors) treated hESC found to inhibit/escape from teratoma Blum et al., 2009 but the mechanism is still to unveil, whereas QC and YM155 treatment blocks Bcl10 and teratoma formation from undifferentiated hESC without affecting lineage-specific differentiation via p53 mediated apoptosis Lee et al., 2013 . Again, CD117+/CD44+ hASC line showed fibroblastoid morphology and expressed both stem (Oct4, Nanog, Sox2) and germ cell (Dazl, Stella) markers Yu et al., 2014 . Bortvin group noticed that Stella, a maternal factor which is not required for germ cell specification in mice Bortvin et al., 2004 . Teratocarcinoma the cell of origin is a germ cell as in the ovaries teratoma is most likely arise from oocytes. But it is unclear whether the transforming event occurs in primordial germ cell or fetal gonocyte. Nanog and Sox2, confirmed the existence of undifferentiated cells in T12-hPSC-derived teratomas. Quantitative RT-PCR showed higher expression of Oct-4 and Nanog in some of the T12-hPSC-derived teratomas Ben-David et al., 2014 . In human seminomas it is found to greater expression of Stellar, Nanog, Gdf3 and Oct4 Clark et al., 2004 and they are in general localized to the ovary and testis. But RA-induced differentiation of hESC expresses Stella. By contrast, hEC expresses Stella at a higher level compared with hESC. Over-expression of Stella does not hamper the stem cell state of hESC, but subsequent RA induction escorts to up regulation of germline- and endodermal-associated genes Wongtrakoongate et al., 2013 . Human ASC most likely originate from primordial germ cell (PGC), and treatment with the HDA inhibitor VPA converted these multipotent cells into pluripotent cell type. Therefore, the transcriptome of hASC is apparently comparable to that of embryonic germ cells (EGC) and PGC. Glioblastoma neural stem cell was reprogrammed by using just two transcription factors Oct4 and Klf4 Stricker and Pollard, 2014 . In presence of O2, glycolysis is enhanced in undifferentiated Oct4-depleted hESC and tumor cells, but OXPHOS is weakened/close up Abu Dawud et al.,2012 . Tributyltin and azadeoxycytidine are the tumor escapers because it induces mitochondrial fission through NAD-IDH dependent mitofusin degradation in human embryonic carcinoma cells and induce apoptosis in mouse ESC Yamada et al., 2015 . In case of hASC, maybe there is an existence of distinct factors in amniotic fluid or site of injection of hASC and their modulations are important for initiation of tumor. As yet, the effects of microenvironments on teratocarcinoma cells have been arrested for extended study.
From the available information, it could be hypothesized that Oct4, Stella and Nanog, Birc5, Blc10, Lrrc50, miR-27 and genes of Wnt and Tgf-β pathways interlinked to form teratocarcinoma within a complex network of unknown mechanism ( Figure 3 ). This network is somehow trafficking by their niches. Hence, it is essential to expose the network of teratoma paradox to exploit regenerative medicine regimes. To prop up the hypothesis, it is needed to study hESC, iPSC, hASC, EGC, PGC and teratoma from stem state to differentiated state followed by embryoid body formation under different culture microenvironment, and sorted out whether there are any overlapping, similar, dissimilar and negatively linked molecular signatures present. Another question is whether the teratoma origin from single hASC or ESC or iPSC or from a multiple cells or what makes it differentiate from other tumor and what gives such cells the ability to form tumors? Knowing the molecular clues of the rising of monster from the angelic cells will be a big catch in regenerative medicine. Needinginvestigation of how comparable hiPSC, hESC, PGC and EGC to hASC in respect to their teratoma formation capabilities upon stimulation/inhibition with distinct small molecules and factors, and thereby, derive a potential hypothesis regarding the mechanism to avoid teratoma formation from hESC and hiPSC. Firstly the approach for the characterization of teratoma reversely from hASC could help the analysis of analogies between teratoma and stem cell pluripotency. By activation/deactivation of specific pathways or the knockdown/over expression of proteins, we may identify the network of transcription factors controlling self-renewal/pluripotency of PSC and ECC. It will give a clue for the cue card “what makes hASC escapable from teratoma in SCID mice”. Further, it will help to find the core candidate for answering the molecular mechanism of teratoma formation upon transplantation, and thereby PSC can be prevented from the tumor.
Conclusion
There is much hope and enthusiasm for stem cell therapy. However, the science still has a lot to catching up to do. On one hand hESC and hiPSC may be differentiated into tissues that could be used in the cell and tissue based therapy of diseased and injured organ. On the other hand, undifferentiated hESC and hiPSC cells persisting in cell grafts have issued of warning as they typically tumorigenic and prone to telomere shortening. To an end, reveal current drawback of stem cells oncogenesis and give a meaningful differences between PSC and hASC from teratoma points of view opens a new horizon for clinical research of stem cell biology.
References
- R. Abu Dawud, K. Schreiber, D. Schomburg, J. Adjaye. Human embryonic stem cells and embryonal carcinoma cells have overlapping and distinct metabolic signatures. PloS one. 2012;7:e39896. Google Scholar
- A.J. Armstrong, M.S. Marengo, S. Oltean, G. Kemeny, R.L. Bitting, J.D. Turnbull, C.I. Herold, P.K. Marcom, D.J. George, M.A. Garcia-Blanco. Circulating tumor cells from patients with advanced prostate and breast cancer display both epithelial and mesenchymal markers. Molecular Cancer Research. 2011;9:997-1007. Google Scholar
- S.G. Basten, E.E. Davis, A.J. Gillis, E. van Rooijen, H. Stoop, N. Babala, I. Logister, Z.G. Heath, T.N. Jonges, N. Katsanis. Mutations in LRRC50 predispose zebrafish and humans to seminomas. PLoS Genet. 2013;9:e1003384. Google Scholar
- U. Ben-David, G. Arad, U. Weissbein, B. Mandefro, A. Maimon, T. Golan-Lev, K. Narwani, A.T. Clark, P.W. Andrews, N. Benvenisty. Aneuploidy induces profound changes in gene expression, proliferation and tumorigenicity of human pluripotent stem cells. Nature communications. 2014;5:. Google Scholar
- B. Blum, O. Bar-Nur, T. Golan-Lev, N. Benvenisty. The anti-apoptotic gene survivin contributes to teratoma formation by human embryonic stem cells. Nature biotechnology. 2009;27:281-287. Google Scholar
- B. Blum, N. Benvenisty. The tumorigenicity of human embryonic stem cells. Advances in cancer research. 2008;100:133-158. Google Scholar
- A. Bortvin, M. Goodheart, M. Liao, D.C. Page. Dppa3/Pgc7/stella is a maternal factor and is not required for germ cell specification in mice. BMC developmental biology. 2004;4:2. Google Scholar
- K.S. Chen, J.F. Amatruda. A big catch for germ cell tumour research. PLoS Genet. 2013;9:e100348-1. Google Scholar
- A.T. Clark, R.T. Rodriguez, M.S. Bodnar, M.J. Abeyta, M.I. Cedars, P.J. Turek, M.T. Firpo, R.A. Reijo Pera. Human STELLAR, NANOG, and GDF3 genes are expressed in pluripotent cells and map to chromosome 12p13, a hotspot for teratocarcinoma. Stem Cells. 2004;22:169-179. Google Scholar
- D. Davydova. Stem cells in human amniotic fluid. Biology bulletin. 2010;37:437-445. Google Scholar
- P. De Coppi, G. Bartsch, M.M. Siddiqui, T. Xu, C.C. Santos, L. Perin, G. Mostoslavsky, A.C. Serre, E.Y. Snyder, J.J. Yoo. Isolation of amniotic stem cell lines with potential for therapy. Nature biotechnology. 2007;25:100-106. Google Scholar
- H. Fuchs, M. Theuser, W. Wruck, J. Adjaye. miR-27 negatively regulates pluripotency-associated genes in human embryonal carcinoma cells. . 2014;:. Google Scholar
- S. Ilancheran, A. Michalska, G. Peh, E.M. Wallace, M. Pera, U. Manuelpillai. Stem cells derived from human fetal membranes display multilineage differentiation potential. Biology of reproduction. 2007;77:577-588. Google Scholar
- C. Koike, K. Zhou, Y. Takeda, M. Fathy, M. Okabe, T. Yoshida, Y. Nakamura, Y. Kato, T. Nikaido. Characterization of amniotic stem cells. Cellular Reprogramming (Formerly “Cloning and Stem Cells”). 2014;16:298-305. Google Scholar
- M.-O. Lee, S.H. Moon, H.-C. Jeong, J.-Y. Yi, T.-H. Lee, S.H. Shim, Y.-H. Rhee, S.-H. Lee, S.-J. Oh, M.-Y. Lee. Inhibition of pluripotent stem cell-derived teratoma formation by small molecules. Proceedings of the National Academy of Sciences. 2013;110:E3281-E3290. Google Scholar
- J. Li, C. Koike-Soko, J. Sugimoto, T. Yoshida, M. Okabe, T. Nikaido. Human Amnion-derived Stem Cells have Immunosuppressive Properties on NK cells and Monocytes. Cell transplantation. 2014;:. Google Scholar
- G.R. Martin, M.J. Evans. The morphology and growth of a pluripotent teratocarcinoma cell line and its derivatives in tissue culture. Cell. 1974;2:163-172. Google Scholar
- D. Moschidou, K. Drews, A. Eddaoudi, J. Adjaye, P. De Coppi, P. V Guillot. Molecular signature of human amniotic fluid stem cells during fetal development. Current stem cell research & therapy. 2013;8:73-81. Google Scholar
- D. Moschidou, S. Mukherjee, M.P. Blundell, K. Drews, G.N. Jones, H. Abdulrazzak, B. Nowakowska, A. Phoolchund, K. Lay, T.S. Ramasamy. Valproic acid confers functional pluripotency to human amniotic fluid stem cells in a transgene-free approach. Molecular Therapy. 2012;20:1953-1967. Google Scholar
- T. Oki, Y. Sowa, T. Hirose, N. Takagaki, R. Nakanishi, M. Kanazawa, Y. Satomi, H. Nishino, T. Miki, T. Sakai. Genistein induces gadd45 gene and G2/M cell cycle arrest in the DU145 human prostate cancer cell line. Cancer Research. 2005;65:574-575. Google Scholar
- M.A. Papadaki, G. Kallergi, Z. Zafeiriou, L. Manouras, P.A. Theodoropoulos, D. Mavroudis, V. Georgoulias, S. Agelaki. Co-expression of putative stemness and epithelial-to-mesenchymal transition markers on single circulating tumour cells from patients with early and metastatic breast cancer. BMC cancer. 2014;14:65-1. Google Scholar
- C.R. Regenbrecht, M. Jung, H. Lehrach, J. Adjaye. The molecular basis of genistein-induced mitotic arrest and exit of self-renewal in embryonal carcinoma and primary cancer cell lines. BMC medical genomics. 2008;1:49. Google Scholar
- S.A. Scherjon, C. Kleijburg-van der Keur, W.A. Noort, F.H. Claas, R. Willemze, W.E. Fibbe, H.H. Kanhai. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood. 2003;102:1548-1549. Google Scholar
- J. Si, J. Dai, J. Zhang, S. Liu, J. Gu, J. Shi, S.G. Shen, L. Guo. Comparative Investigation of Human Amniotic Epithelial Cells and Mesenchymal Stem Cells for Application in Bone Tissue Engineering. Stem cells international 2015. 2015;:. Google Scholar
- S. Stricker, S. Pollard. Reprogramming cancer cells to pluripotency: an experimental tool for exploring cancer epigenetics. Epigenetics. 2014;9:798-802. Google Scholar
- L. Sun, R. Gong, B. Wan, X. Huang, C. Wu, X. Zhang, S. Zhao, L. Yu. GADD45γ, down-regulated in 65% hepatocellular carcinoma (HCC) from 23 Chinese patients, inhibits cell growth and induces cell cycle G2/M arrest for hepatoma Hep-G2 cell lines. Molecular biology reports. 2003;30:249-253. Google Scholar
- B. Sung, H.-J. Do, S.-W. Park, S.-H. Huh, J.-H. Oh, H.-J. Chung, M.-J. Kang, J.-H. Kim, N.-H. Kim, J.-H. Kim. Regulation of OCT4 gene expression by liver receptor homolog-1 in human embryonic carcinoma cells. Biochemical and biophysical research communications. 2012;427:315-320. Google Scholar
- J.A. Wakeman, J. Walsh, P.W. Andrews. Human Wnt-13 is developmentally regulated during the differentiation of NTERA-2 pluripotent human embryonal carcinoma cells. Oncogene. 1998;17:179-186. Google Scholar
- P. Wongtrakoongate, M. Jones, P.J. Gokhale, P.W. Andrews. STELLA facilitates differentiation of germ cell and endodermal lineages of human embryonic stem cells. PloS one. 2013;8:. Google Scholar
- S. Yamada, Y. Kotake, M. Nakano, Y. Sekino, Y. Kanda. Tributyltin induces mitochondrial fission through NAD-IDH dependent mitofusin degradation in human embryonic carcinoma cells. Metallomics. 2015;:. Google Scholar
- X. Yu, N. Wang, R. Qiang, Q. Wan, M. Qin, S. Chen, H. Wang. Human amniotic fluid stem cells possess the potential to differentiate into primordial follicle oocytes in vitro. Biology of reproduction, biolreprod. 113.112920. 2014;:. Google Scholar



![[Download figure]](s40855-015-0002-1/fig1.png){kind=link}
![[Download figure]](s40855-015-0002-1/fig2.png){kind=link}
![[Download figure]](s40855-015-0002-1/fig3.png){kind=link}
(a) Flow cytometry plot of unstained ASC confirmed the presence of both small (circled in red) and larger (circled in blue) granular cells. On the right, representative phase-contrast image of hASC in suspension showing the presence of small (S) and larger (L) cells. (b) Alkaline phosphatase staining of hASC. C) Immunostained stem cell marker expressed in hASC. Figure adapted from Moschidou et al. (2012).